Finalmente è uscito il Numero 8 di Fossil&Minerals Review. In questo numero andremo indietro nel tempo fino al Carnico per un viaggio sulle barriere coralline di 230 milioni di anni fa per poi spostarci in un paradiso per i micromont, ovvero le cave di Cuasso al Monte in provincia di Varese.
In questi mesi due pilastri della paleontologia del Centro Italia ci hanno lasciato. Un breve ricordo per entrambi da parte della redazione della rivista.
Buona Lettura!
Indice
Ricerche Mineralogiche a Cuasso al Monte, Varese
di Stefano Mercadante pag.1
Ricordo del Prof. Federico Venturi
di Federico Famiani pag. 27
Serafino Calindri e i fossili del bolognese
di Romano Guerra pag.28
Il Livello Faraoni
di Paolo Faraoni pag. 39
Bigiaretti, Addio!
di Romano Guerra pag. 49
Formazione di San Cassiano:
barriere coralline del Carnico a 2000 metri
di Claudio Zuccaro Pag.50







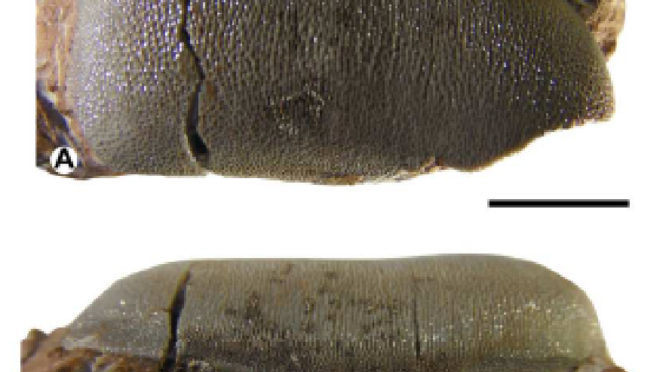
 presenta il primo ritrovamento riferibile allo squalo durofago nella Formazione Rosso Ammonitico del Toarciano (Giurassico inferiore). Sono stati recuperati Due denti isolati che erano conservati all’interno della marna calcarea del Rosso ammonitico Toarciano (zona dei bifrons). Il dente meglio conservato è caratterizzato da una struttura generale che ben si accorda con la classica dentatura schiacciata tipica degli squali durofagi della sottofamiglia Acrodontinae. I caratteri morfologici analizzati nel lavoro consentono di attribuire il materiale rinvenuto al genere Asteracanthus sp.
presenta il primo ritrovamento riferibile allo squalo durofago nella Formazione Rosso Ammonitico del Toarciano (Giurassico inferiore). Sono stati recuperati Due denti isolati che erano conservati all’interno della marna calcarea del Rosso ammonitico Toarciano (zona dei bifrons). Il dente meglio conservato è caratterizzato da una struttura generale che ben si accorda con la classica dentatura schiacciata tipica degli squali durofagi della sottofamiglia Acrodontinae. I caratteri morfologici analizzati nel lavoro consentono di attribuire il materiale rinvenuto al genere Asteracanthus sp.